Welcome to the Hunter lab
The goal of the Hunter Lab is to understand the molecular basis of cell growth control and cell cycle regulation. Many growth factor receptors are protein-tyrosine kinases (PTK), which are activated by ligand-induced dimerization. Mitogenic signalling by PTKs involves tyrosine phosphorylation of critical target proteins. We are currently investigating what signal pathways are activated by the mammalian PDGF and stem call factor receptor PTKs, and the EphA2 receptor PTK, and investigating the roles of the Nck and c-Cbl adaptor proteins that are phosphorylated by activated receptor PTKs. We are also studying what signalling pathways are stimulated when integrin receptors bind to extracellular matrix proteins, such as fibronectin, in mammalian cells. We are focusing on the roles of the focal adhesion kinase (FAK) nonreceptor PTK and its relative Pyk2, and the c-Src family of nonreceptor PTKs, which are activated upon adhesion. We have also identified a Drosophila FAK/Pyk2 homologue, DmFAK, and we are carrying out a genetic analysis of its function.
To complement our work on PTKs, we are investigating the functions of protein-tyrosine phosphatases (PTPs) in cell signalling. We are studying the receptor-like PTP, RPTPa, which like most RPTPs has a twin catalytic domain structure. The crystal structure of the membrane proximal catalytic domain of RPTPa shows that it exists as an inactive dimer, suggesting that dimerization of RPTPa, and other RPTPs, may negatively regulate their activity. We are have shown that this is true for the CD45 RPTP, and have evidence that RPTPa activity is also inhibited by dimerization. This principle of regulation would be the exact opposite of that observed for receptor PTKs.
Our studies on the role of phosphorylation in mammalian cell cycle regulation are focused on the cyclin-dependent kinases (Cdk), and their substrates and inhibitors, and on the Cdc7/Dbf4 kinase. Using a new protein kinase substrate screen, we have identified Prc1, a spindle-associated protein, as a Cdk substrate, and we have also shown that Cdc6, an essential component of the prereplication complex that is bound to chromosomal origins, is phosphorylated by cyclin E/Cdk2, resulting in its export from the nucleus thus preventing rereplication. We have identified a novel peptidyl-prolyl isomerase, Pin1, that contains a WW domain and a prolyl isomerase domain. Pin1 is the functional homologue of yeast Ess1p, a protein essential for progression through mitosis. We have solved the structure of Pin1, and shown that it binds to phosphorylated Ser.Pro sequences, which are generated by Cdk phosphorylation. We are studying the role of Pin1 in cell cycle progression.
News Highlights

Celebrating 50 years of discovery: Professor Tony Hunter’s half-century legacy at the Salk Institute
LA JOLLA—The Salk Institute is proud to celebrate Professor Tony Hunter’s 50 years as a cancer biology pioneer whose fundamental discoveries have inspired the development of more than 80 cancer drugs. Since joining the Institute’s faculty in February 1975, Hunter has been a cornerstone of the Salk community, contributing to transformational discoveries and mentoring more than 100 trainees, many of whom have also become scientific leaders.
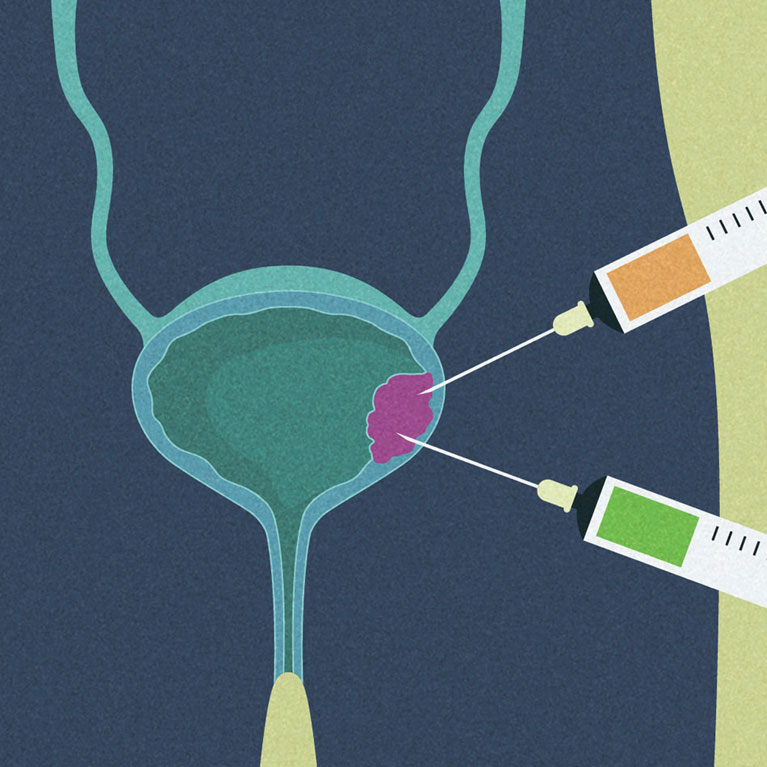
Putting a lid on excess cholesterol to halt bladder cancer cell growth
LA JOLLA—Like all cancers, bladder cancer develops when abnormal cells start to multiply out of control. But what if we could put a lid on their growth?

Salk Institute celebrates 50th anniversary and renewal of National Cancer Institute designation
LA JOLLA—The Salk Institute marks 50 years as a National Cancer Institute (NCI)-Designated Cancer Center with good news: NCI has renewed the designation and grant support for another five years.

Three Salk scientists among 2022 Curebound Discovery Grant winners
LA JOLLA—The Salk Institute’s American Cancer Society Professor Tony Hunter, Professor Reuben Shaw, and Assistant Professor Graham McVicker are among 12 inaugural 2022 Discovery Grant winners. The awards, which total $3 million, were launched this year by Curebound, a philanthropic organization dedicated to funding collaborative cancer research that has the potential to reach the clinic.

Salk scientist Tony Hunter receives 2022 AACR Lifetime Achievement Award in Cancer Research
LA JOLLA—Salk Institute Professor Tony Hunter will receive the 2022 American Association for Cancer Research (AACR) Award for Lifetime Achievement in Cancer Research at the April annual meeting of AACR, the largest cancer research organization in the world dedicated to preventing and curing all cancers. This major award is a significant recognition of Hunter’s contributions to cancer research, which have led to the development of the highly effective leukemia drug GleevecTM.

The Lustgarten Foundation and Salk Institute announce strategic pancreatic cancer research partnership
LA JOLLA/NEW YORK—The Lustgarten Foundation and Salk Institute today announced a new strategic partnership supported by a $5 million grant and focused on identifying and validating potential targets for new pancreatic cancer drugs. The effort will be led by four co-principal investigators, all prominent cancer researchers in the Salk Dedicated Program in Pancreatic Cancer: Professors Reuben Shaw, Ronald Evans, Tony Hunter and Assistant Professor Dannielle Engle. The partnership is part of the Lustgarten Advancing Breakthrough Science (LABS) Program.
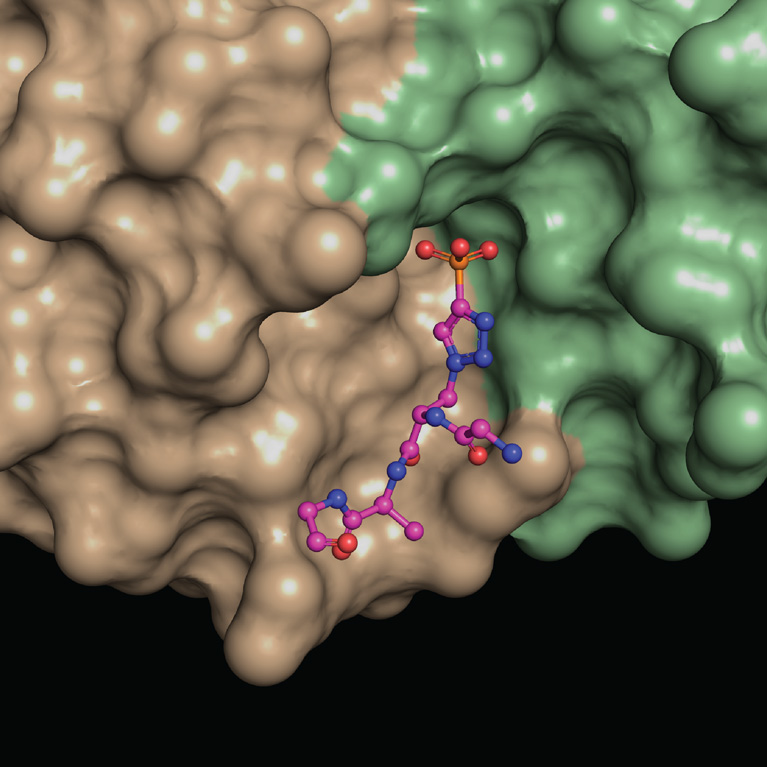
Salk team reveals never-before-seen antibody binding, informing both liver cancer and antibody design
LA JOLLA—In structural biology, some molecules are so unusual they can only be captured with a unique set of tools. That’s precisely how a multi-institutional research team led by Salk scientists defined how antibodies can recognize a compound called phosphohistidine—a highly unstable molecule that has been found to play a central role in some forms of cancer, such as liver and breast cancer and neuroblastoma.

Salk researchers accelerate, expand COVID-19 research
LA JOLLA—As the COVID-19 pandemic continues across the globe, the Salk Institute joins in efforts to understand the fundamental science behind the novel coronavirus to pave the way to treatments and cures. COVID-19 exploits a vulnerability in the immune system’s armor: because the SARS-CoV-2 virus—the novel coronavirus that causes COVID-19—appeared in humans recently, our immune systems have no experience with the virus—and sometimes have difficulty fighting it.
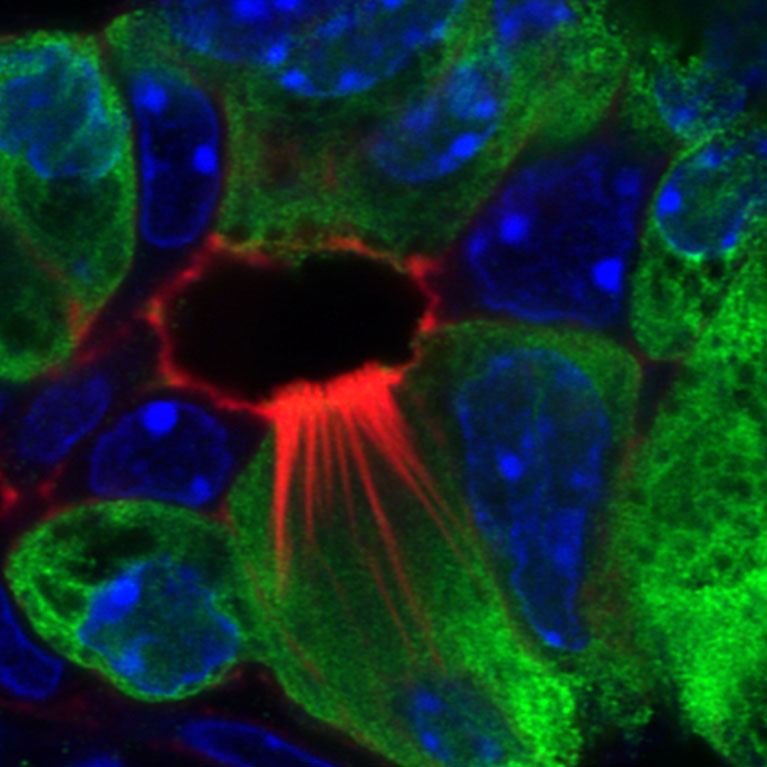
Mysterious tuft cells found to play role in pancreatitis
LA JOLLA—Persistent inflammation of the pancreas (chronic pancreatitis) is a known risk factor for developing pancreatic cancer, the third-deadliest cancer in the United States. Tuft cells—cells sensitive to chemical (chemosensory) changes typically found in the intestines and respiratory tract—had previously been discovered in the pancreas, but their function has largely remained a mystery. Now, a team of Salk scientists led by Professor Geoffrey Wahl and Staff Scientist Kathleen DelGiorno has uncovered the formation of tuft cells during pancreatitis and the surprising role of tuft cells in immunity, using mouse models of pancreatitis. The findings, published in Frontiers in Physiology on February 14, 2020, could lead to the development of new biomarkers to test for pancreatitis and pancreatic cancer.

Salk Institute hits play on new podcast series
LA JOLLA—A new podcast series called Where Cures Begin launches this week and features one-on-one conversations with Salk researchers working at the forefront of their respective scientific fields, from cancer and neuroscience to plant biology, circadian science and more. The eight episodes of season 1, which will be released weekly beginning October 30, 2019, include interviews with the following Salk scientists:

Salk scientist Tony Hunter receives National Cancer Institute Outstanding Investigator Award
LA JOLLA, CA—Salk scientist Tony Hunter has received a National Cancer Institute (NCI) Outstanding Investigator Award (OIA), which supports accomplished leaders in cancer research. Hunter, who is an American Cancer Society Professor, will receive more than $7,500,000 over the next seven years to further his work. According to the NCI, the award supports investigators who are providing significant contributions toward understanding cancer and developing applications that may lead to a breakthrough in cancer research.
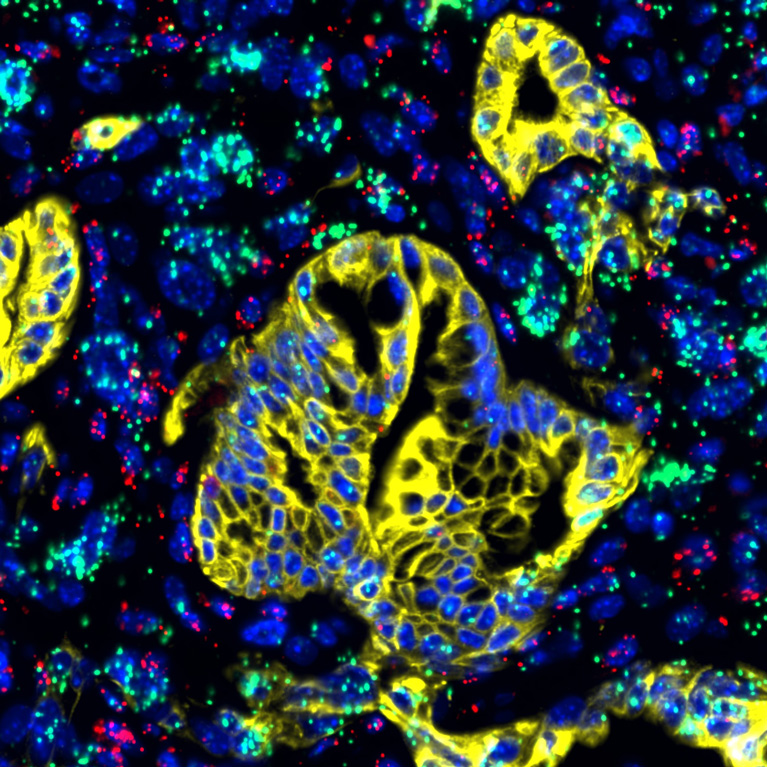
New study targets Achilles’ heel of pancreatic cancer, with promising results
LA JOLLA—Advanced pancreatic cancer is often symptomless, leading to late diagnosis only after metastases have spread throughout the body. Additionally, tumor cells are encased in a “protective shield,” a microenvironment conferring resistance to many cancer treatment drugs. Now, Salk Institute researchers, along with an international team of collaborators, have uncovered the role of a signaling protein that may be the Achilles’ heel of pancreatic cancer.

Research in yeast leads to serendipitous finding about a central nervous system disorder
LA JOLLA—Studying the fundamental aspects of biology can sometimes lead to unexpected findings that directly relate to human disease. In one of the latest examples of scientific serendipity, researchers from the Salk Institute found that an important quality control mechanism in baker’s yeast is closely connected to hypomyelinating leukodystrophy, a debilitating disease found in children.

What the satellites in your body do
LA JOLLA—Only some of us have satellite TV in our homes, but all of us have satellite DNA in cells in our bodies. Working copies of satellite DNA (called satellite RNAs) are high in certain types of cancer, such as breast and ovarian. But whether they cause cancer or merely coincide with it has been unclear.
Salk researchers win five competitive grants for cancer research
LA JOLLA—Salk Institute scientists Ronald Evans, Diana Hargreaves, Tony Hunter, Graham McVicker and Geoffrey Wahl are among the first wave of researchers to receive funding from Padres Pedal the Cause, one of one of the largest stand-alone cancer fundraising events in San Diego. The nonprofit raised $2.4 million for cancer research in November 2017, thanks to the efforts of more than 3,000 bicycle riders, sponsors, volunteers and donors.

Salk Institute’s Tony Hunter wins 2018 Tang Prize in Biopharmaceutical Science
LA JOLLA—Salk American Cancer Society Professor Tony Hunter has been awarded the 2018 Tang Prize in Biopharmaceutical Science.

Tumor suppressor protein targets liver cancer
LA JOLLA—Salk Institute scientists, together with researchers from Switzerland’s University of Basel and University Hospital Basel, discovered a protein called LHPP that acts as a molecular switch to turn off the uncontrolled growth of cells in liver cancer. The tumor suppressor, which could be useful as a biomarker to help diagnose and monitor treatment for liver cancer, could also be relevant for other cancer types. The work appeared in print in the journal Nature on March 29, 2018, and adds to the growing body of knowledge about cellular processes that either promote or prevent cancer.

Tony Hunter receives Pezcoller Foundation–AACR International Award for Extraordinary Achievement in Cancer Research
LA JOLLA—Professor Tony Hunter, who holds an American Cancer Society Professorship at the Salk Institute, has received the 2018 Pezcoller–AACR International Award for Extraordinary Achievement in Cancer Research, one of the most prestigious honors in the field of cancer research. The prize recognizes a scientist of international renown who has made a major scientific discovery in basic cancer research or who has made significant contributions to translational cancer research.

Salk scientist awarded inaugural Sjöberg Prize for cancer breakthrough
LA JOLLA—Salk Professor Tony Hunter, who holds an American Cancer Society Professorship, has been awarded $500,000 as part of the $1 million Royal Swedish Academy of Sciences’ inaugural Sjöberg Prize for Cancer Research for “groundbreaking studies of cellular processes that have led to the development of new and effective cancer drugs.” The prize ceremony, which is modeled after the Nobel Prize ceremony, will be held in Stockholm during the Academy’s annual meeting on March 31, 2017, in the presence of His Majesty the King and Her Majesty the Queen of Sweden.
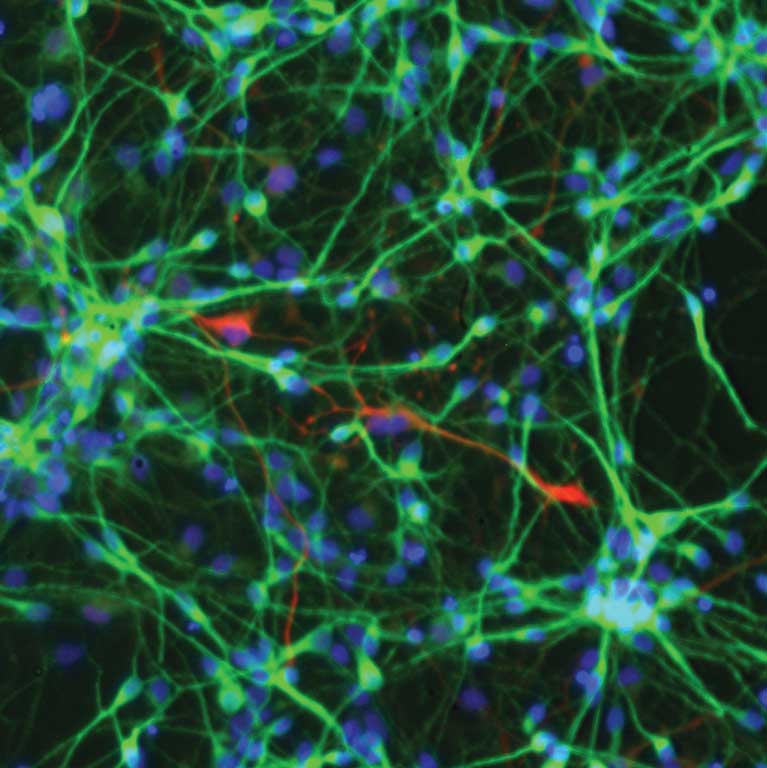
Power up: growing neurons undergo major metabolic shift
LA JOLLA—Our brains can survive only for a few minutes without oxygen. Salk Institute researchers have now identified the timing of a dramatic metabolic shift in developing neurons, which makes them become dependent on oxygen as a source of energy.
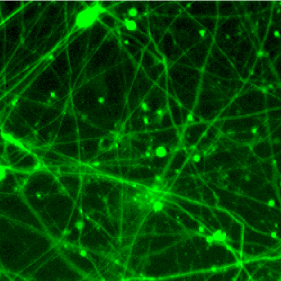
Tamping down neurons’ energy use could treat neurodegeneration
LA JOLLA—Salk Institute scientists showed how an FDA-approved drug boosts the health of brain cells by limiting their energy use. Like removing unnecessary lighting from a financially strapped household to save on electricity bills, the drug—called rapamycin—prolongs the survival of diseased neurons by forcing them to reduce protein production to conserve cellular energy.

New technique maps elusive chemical markers on proteins
LA JOLLA–Unveiling how the 20,000 or so proteins in the human body work–and malfunction–is the key to understanding much of health and disease. Now, Salk researchers developed a new technique that allows scientists to better understand an elusive step critical in protein formation.
Tony Hunter wins BBVA Foundation Frontiers of Knowledge Award in Biomedicine
LA JOLLA–Tony Hunter, professor and director of the Salk Institute Cancer Center, in La Jolla, California, has received the BBVA Foundation Frontiers of Knowledge Award in the Biomedicine category for “carving out the path that led to the development of a new class of successful cancer drugs.”
Salk researcher Tony Hunter to receive 2014 Royal Medal in biological sciences
LA JOLLA–Salk Institute professor Tony Hunter has been awarded the 2014 Royal Medal for biological sciences by the Royal Society, a fellowship of some of the world’s most eminent scientists based in the United Kingdom.

Pedal the Cause funds local ground-breaking cancer research with inaugural grant awards
LA JOLLA—Pedal the Cause, the region’s only multiday cycling fundraiser where 100 percent of the net proceeds goes to support cancer research in San Diego, today announced five research projects funded by the inaugural 2013 event. The Pedal the Cause grants offer enough support for initial experiments, which will ideally lead to grants from federal sources and to large-scale studies.

American Association for Cancer Research appoints Salk scientists to inaugural class of fellows
LA JOLLA, CA—The American Association for Cancer Research (AACR), the world’s oldest and largest professional organization dedicated to accelerating scientific progress to prevent and cure cancer, has selected four Salk scientists and two of the Institute’s nonresident fellows to be inducted in its first class of the fellows of the AACR Academy.

What can the water monster teach us about tissue regeneration in humans?
LA JOLLA, CA—Based on two new studies by researchers at the Salk Institute for Biological Studies, regeneration of a new limb or organ in a human will be much more difficult than the mad scientist and supervillain, Dr. Curt Connors, made it seem in the Amazing Spider-man comics and films.
Salk Institute hosts Cell Cycle Symposium – Discussions that change lives
La Jolla—The 2011 Cell Cycle meeting will be held at the Salk Institute for Biological Studies in La Jolla, California, starting at 7:00 P.M. on Friday, June 17 and ending on Monday, June 20, 2011. Since 1999 the Salk Institute has held the Cell Cycle Meeting every other year, alternating with Cold Spring Harbor, where the Cell Cycle Meeting was initiated in 1992.
Salk Institute cancer researcher Tony Hunter named to Frederick W. and Joanna J. Mitchell Chair
LA JOLLA, CA—The Salk Institute for Biological Studies today announced the appointment of Professor Tony Hunter, Ph.D., as the inaugural holder of the Frederick W. and Joanna J. Mitchell Chair, created in memory of their daughter Marian Mitchell through a $2 million gift by the estate of Frederick W. Mitchell.
Mobilizing the repair squad: Critical protein helps mend damaged DNA
LA JOLLA, CA—In order to preserve our DNA, cells have developed an intricate system for monitoring and repairing DNA damage. Yet precisely how the initial damage signal is converted into a repair response remains unclear. Researchers at the Salk Institute for Biological Studies have now solved a crucial piece of the complex puzzle.
Chemotherapy resistance: Checkpoint protein provides armor against cancer drugs
LA JOLLA, CA-Cell cycle checkpoints act like molecular tripwires for damaged cells, forcing them to pause and take stock. Leave the tripwire in place for too long, though, and cancer cells will press on regardless, making them resistant to the lethal effects of certain types of chemotherapy, according to researchers at the Salk Institute for Biological Studies.
Climbing the ladder to longevity: critical enzyme pair identified
LA JOLLA, CA-Experiment after experiment confirms that a diet on the brink of starvation expands lifespan in mice and many other species. But the molecular mechanism that links nutrition and survival is still poorly understood. Now, researchers at the Salk Institute for Biological Studies have identified a pivotal role for two enzymes that work together to determine the health benefits of diet restriction.

Breast cancer cells have to learn to walk before they can run
La Jolla, CA – Early stage breast cancer that has not yet invaded the surrounding tissues may already contain highly motile cells, which brings the tumor one step closer to metastasis, report researchers at the Salk Institute for Biological Studies.
Get in Touch First
La Jolla, CA – When the genetic material inside a cell’s nucleus starts to fall apart, a protein called ATM takes charge and orchestrates the rescue mission. Surprisingly, for ATM to kick into full gear, the stretches of DNA flanking a chromosomal break are just as important as the damaged site itself, report scientists at the Salk Institute for Biological Studies.
Tony Hunter receives Robert. J. and Claire Pasarow Award for Cancer Research
La Jolla, CA – Salk scientist Dr. Tony Hunter is the recipient of the 2006 Pasarow Award in Cancer Research for his key discoveries of the chemical “switch” that turns healthy cells into cancer cells.
Distinguishing friend from foe in the battle against cancer
La Jolla, CA – The latest generation of cancer chemotherapeutic drugs specifically targets mutant enzymes or “oncoproteins” that have run amok and now promote uncontrolled cell growth. As promising as these drugs are, cancer cells with their backs against the wall have the tendency to fight back. A major goal of cancer research is to frustrate these acts of cellular desperation.
Genetically modified mice are resistant to obesity despite a high fat diet
La Jolla, CA – Ravenous mice that chomp down as if there were no tomorrow yet stay lean and mean? Shutting down two genes that modulate a body’s energy balance transformed these animals into fidgeting, highly efficient fat burning machines, report scientists at the Salk Institute for Biological Studies in this week’s issue of Cell Metabolism.
Molecular ‘zipcode’ guides nerves to correct places in body
La Jolla, CA – During embryonic development, thousands of nerves must be connected to muscles as part of a communication network that allows the newborn to move, breathe and lead a normal life. The question is, how does this complicated ‘telephone system’ get wired up?
Salk Scientist who discovered cancer ‘switch’ awarded the Wolf Prize in Medicine
La Jolla, CA – Salk Institute scientist Tony Hunter has been awarded the 2005 Wolf Prize in Medicine, Israel’s top recognition for achievements in the interest of mankind, for his key discoveries in cell regulation and cancer research.
Salk Scientist Who Revolutionized Cancer Research Awarded Horwitz Prize
La Jolla, CA – Salk Institute scientist Tony Hunter has been awarded the 2004 Louisa Gross Horwitz Prize, a leading national award for scientific achievement, for his pivotal discoveries about the chemical ‘switch’ that turns healthy cells into cancer cells.
Salk Institute Scientist Tony Hunter Named to Institute of Medicine
La Jolla, CA – Salk Institute for Biological Studies scientist Tony Hunter has been named a member of the prestigious Institute of Medicine (IOM) of the National Academies of Sciences, Dr. Richard Murphy, Salk Institute President said today.
Hunter Receives Two Major Cancer Awards
La Jolla, CA – Tony Hunter, professor of molecular and cell biology at the Salk Institute, has received two major awards for cancer research.
Salk Researchers Ranked in Top 25 of Scientific Citations Worldwide
La Jolla, CA – Two Salk Institute researchers ranked within the top 25 scientists worldwide for their work’s influence in any field, according to an organization that monitors the impact of scientific publications.
Missing Enzyme Found to Lead to Alzheimer?s Disease Symptoms
La Jolla, CA – An enzyme studied by scientists for years can prevent tangles of proteins from forming in the brain’s nerve cells – as occurs in patients with age-dependent Alzheimer’s disease, according to a study by investigators at the Salk Institute, Beth Israel Deaconess Medical Center and Harvard Medical School.
Salk Institute and SUGEN Scientists Map ‘Human Kinome’
La Jolla, CA – A California research team has mapped an entire group of human enzymes, providing important information for the development of a new generation of drugs to treat cancer and other diseases. The findings will be published in the Dec. 6 issue of Science.
Potential Male Contraceptive Target Discovered by Salk Scientists
La Jolla, CA – The first gene to control male fertility without affecting sexual behavior or physical appearance has been identified by Salk researchers. The study, appearing in the current Nature Genetics, demonstrates that a certain genetic mutation eliminates sperm development in male mice. It has no effect on females.